№«ЛҫөШЦ·ЈәДПҫ©КРҫӯјГјјКхҝӘ·ўЗшәгҫәВ·31-1әЕ
өз»°Јә025Јӯ86581148


1 ЬҝМҰЛШДЪхҘКЗөЪБщҙуЦІОпјӨЛШ
ЎЎЎЎЦІОпФЪЙПТЪДкөДҪш»Ҝ№эіМЦРЦрҪҘІъЙъТ»МЧЧФОТөчҪЪПөНіТФУҰ¶ФёчЦЦ»·ҫіРІЖИЈ¬¶шХвЦЦөчҪЪПөНіНЁіЈКЗУЙЦІОпДЪФҙјӨЛШРЕәЕөДҙ«өјУлПмУҰАҙҝШЦЖөДЎЈХвР©ОўБҝөДРЎ·ЦЧУУР»ъ»ҜәПОпРӯөчҝШЦЖЦІОпөДЙъіӨ·ўУэј°ДжҫіРІЖИПмУҰЎЈЛжЧЕИЛАа¶ФЦІОпјӨЛШөДЙоИлБЛҪвЈ¬ҝӘКјҪ«МмИ»ЦІОпјӨЛШ»тИЛ№ӨәПіЙјӨЛШАаЛЖОпЧчОӘЦІОпЙъіӨөчҪЪјБФЛУГФЪЕ©КВЙъІъЈ¬МбёЯЧчОпІъБҝЎўФцЗҝЧчОпҝ№РФЎўМбЙэЧчОпЖ·ЦКТФВъЧгИЛГЗИХТжЖИЗРөД¶ФЕ©ЧчОпІъБҝј°Ж·ЦКөДТӘЗуЎЈ
ЎЎЎЎФЪЖ·АаЦЪ¶аөДЦІОпЙъіӨөчҪЪјБЦРЈ¬ЬҝМҰЛШДЪхҘЈЁbrassinolideЈ¬BLЈ©ОЮТЙКЗЖдЦРөДЩ®Щ®ХЯЈ¬ЧФ·ўПЦЦ®іхҫНіЙОӘҝЖС§јТИИТйөД»°МвәНЕ©Тө№ӨЧчХЯөДіи¶щЎЈ
ЎЎЎЎГА№ъҝЖС§јТMitchellУЪ1970ДкКЧПИМбіцУНІЛЛШЈЁbrassinsЈ©өДёЕДоЈ¬ХвЦЦАҙФҙУЪУНІЛ»Ё·ЫМбИЎОпөДОҙЦӘіЙ·ЦФЪј«өНөДЕЁ¶ИПВҫНДЬ№»УРР§ҙЩҪшҫҘЙміӨәНПё°ы·ЦБСЎЈЛжәуЈ¬1979ДкҝЖС§јТGroveҙУУНІЛ»Ё·ЫЦРМбИЎөҪТ»ёцҫЯУРУНІЛЛШ»оРФөД»ҜәПОпІўНЁ№эX-ЙдПЯ·ЦОцИ·¶ЁЖд»ҜС§Ҫб№№КҪЈ¬ЧоЦХГьГыОӘЬҝМҰЛШДЪхҘЈЁbrassinolideЈ¬BLЈ©Ј¬УЙУЪЬҝМҰЛШДЪхҘКЗЦЪ¶аУНІЛұніІҙјАаОпЦКЈЁBrassinosteroidЈ¬BRЈ©Ҫб№№ЦР»оРФЧоёЯөДТ»ёцЈ¬ҙЛәуФтК№УГЬҝМҰЛШДЪхҘЧчОӘёГАајӨЛШөДҙъіЖЎЈЦұөҪ1998ДкөЪ16Ҫм№ъјКЦІОпЙъіӨОпЦКС§»бДк»бЙПЈ¬BLІЕұ»ХэКҪИ·ИПОӘөЪБщҙуЦІОпјӨЛШЎЈ
ЎЎЎЎBL№г·әҙжФЪУЪёчЦЦЦІОпМеДЪЈ¬ҙУөНөИөДФеАаЎўЮ§АаЈ¬өҪВгЧУЦІОпЎўұ»ЧУЦІОпЦРТС·ўПЦјёК®ЦЦІ»Н¬Ҫб№№өДЬҝМҰЛШДЪхҘЈ¬ЛөГчBLөДРЕәЕНҫҫ¶ІъЙъУЪЦІОпҪш»ҜөДФзЖЪЈ¬ЗТФЪЦІОпЦРПа¶ФұЈКШЎЈBLФЪЦІОпМеДЪЦчТӘјҜЦРФЪЙъЦіЖч№ЩЎўОҙіЙКмЦЦЧУј°УЧДЫЧйЦҜЦРЈ¬өчҝШЙъЦі·ўУэЎўЦЦЧУіЙКмЈ¬ҙЩҪшҫҘөДЙміӨј°ёщРОМ¬ҪЁіЙЈ¬Н¬КұФЪЦІОпҝ№ДжҫіРІЖИЦРЖрөҪ»эј«ЧчУГЎЈ
2 ЬҝМҰЛШДЪхҘөДМеДЪәПіЙҙъР»УлРЕәЕПмУҰ
2.1 ЬҝМҰЛШДЪхҘөД»ҜС§Ҫб№№
ЎЎЎЎBRКЗТ»АаЦІОпзЮҙјјӨЛШЈ¬Жд»ҜәПОпөД»щұҫҪб№№КЗ·ЦұрУЙABCDЛД»·ЧйіЙөДзЮМеәЛј°B»·ЙПC6О»өДНӘ»щәНD»·C17О»өДНй»щІаБҙЎЈA»·әННй»щІаБҙөДфЗ»щ»ҜЛ®ЖҪј°ҝХјдҪб№№ЦұҪУУ°ПмЬҝМҰЛШДЪхҘУлЖдКЬМеёҙәПОпөДҪбәПЈ¬ҙУ¶шУ°ПмЖдЙъОп»оРФЈ¬МмИ»ЬҝМҰЛШДЪхҘBLКЗДҝЗ°ТСЦӘ»оРФЧоёЯЎўСРҫҝЧоНёі№өДЬҝМҰЛШДЪхҘЈ¬ИЛГЗТСёщҫЭЖдҪб№№КҪЈЁНј1Ј©ОӘА¶ұҫЈ¬ИЛ№ӨәПіЙБЛ24-ұнЬҝМҰЛШДЪхҘЎў28-ёЯЬҝМҰЛШДЪхҘЎў28-ұнёЯЬҝМҰЛШДЪхҘЎў»мұнЬҝМҰЛШДЪхҘөИ¶аЦЦАаЛЖОпЈ¬Іў№г·әУҰУГУЪЕ©ТөЙъІъЎЈ
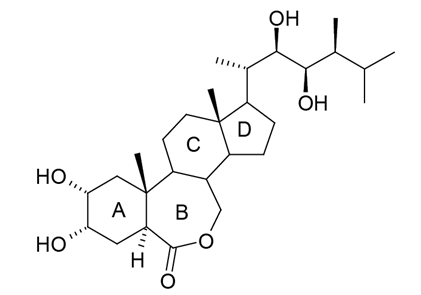
Нј1 МмИ»ЬҝМҰЛШДЪхҘҪб№№КҪ
2.2 ЬҝМҰЛШДЪхҘөДәПіЙҙъР»
ЎЎЎЎФзЖЪЈ¬ҝЖС§јТАыУГіӨҙә»ЁЈЁCatharanthus roseusЈ©РьёЎПё°ыәНН¬О»ЛШКҫЧЩ·ЁСРҫҝBLөДМеДЪәПіЙНҫҫ¶Ј¬И·¶ЁІЛУНзЮҙјЈЁcampesterolЈ©ОӘBRМеДЪәПіЙЗ°МеОпЦКЈ¬Ҫш¶шУЦ·ўПЦІизЮНӘЎўПгЖСзЮҙјәНұНВйзЮНӘөИЦРјдІъОпЈ¬ҫӯАъТ»ПөБРГёҙЩ·ҙУҰЧоЦХРОіЙЬҝМҰЛШДЪхҘЎЈ
ЎЎЎЎҙУІЛУНзЮҙјәПіЙөДөЪТ»ІҪКЗУЙзЮҙј»№ФӯГёDET2ҙЯ»ҜЙъіЙІЛУНзЮНйҙјЈЁcampestanolЈ©ЎЈФЪ¶ФBRәПіЙҙъР»ЙоИлСРҫҝ·ўПЦЈ¬ЬҝМҰЛШДЪхҘC6О»өДјУСхІҪЦиҝЙ·ўЙъФЪәПіЙНҫҫ¶ФзЖЪЈЁФзЖЪC6јУСхНҫҫ¶Ј©Ј¬ТІҝЙ·ўЙъФЪәПіЙНҫҫ¶НнЖЪЈЁНнЖЪC6јУСхНҫҫ¶Ј©Ј¬ХвБҪЦЦәПіЙНҫҫ¶ФЪЦІОпМеДЪКЗІўҙжөДЎЈBLНй»щІаБҙC22О»өДфЗ»щ»Ҝ·ҙУҰКЗәПіЙ№эіМөДПЮЛЩІҪЦиЈ¬УЙCYP90 P450өҘјУСхГёDWF4ҙЯ»ҜНкіЙЈ¬ТтҙЛDWF4КЗBLәПіЙөДПЮЛЩГёЎЈНЁіЈЈ¬BRәПіЙН»ұдМеЈ¬Изdet2әНdwf4Ј¬¶јПФКҫіцөдРНөДBRИұК§Н»ұдМеұнРНЎӘЎӘј«¶И°«»ҜЈ¬ј°ЖдЛы·ўУэТміЈұнРНЎЈ
2.3 Пё°ыДЪBRРЕәЕПмУҰ
ЎЎЎЎBRөДМеДЪәПіЙұ»ИПОӘ·ўЙъФЪПё°ыДЪЦКНшЈ¬ФЩҫӯёЯ¶ы»щМе·ЦГЪЦБ°ыНвЈ¬ФЪР§УҰІҝО»өДПё°ыДӨЙПBRұ»КЬМеҪбәПЈ¬ҙУ¶шјӨ»оПё°ыДЪөДТ»ПөБРРЕәЕҙ«өјНҫҫ¶ЎЈ
ЎЎЎЎЈЁ1Ј©ОЮјӨЛШМхјюПВBRРЕәЕТЦЦЖЎЈПё°ыДӨЙПҙжФЪBRКЬМеө°°ЧBRI1Ј¬ТФј°КЬМеёҙәПОпө°°ЧBAK1ЎЈөұBRІ»ҙжФЪКұЈ¬BRI1әНBAK1·ЦұрУлЖдТЦЦЖө°°ЧBKI1әНBIR3Па»ҘЧчУГЈ¬ЧиЦ№BRКЬМеёҙәПОпөДРОіЙЈ¬К№BRРЕәЕНЁВ·ҙҰУЪ№ШұХЧҙМ¬ЎЈПё°ыДЪө°°ЧјӨГёBIN2КЗBRРЕәЕөДёәөчҝШТтЧУЈ¬ҝЙК№BRЧӘВјТтЧУBES1әНBZR1БЧЛб»Ҝ¶шЦНБфФЪПё°ыЦКЈ¬ОЮ·ЁҪшИлПё°ыәЛРРК№ЧӘВјТтЧУ№ҰДЬЎЈ
ЎЎЎЎЈЁ2Ј©BRКЬМеёҙәПОпјӨ»оЎЈөұBRҙжФЪКұЈ¬КЬМеө°°ЧBRI1°ыНвҪб№№ҝЙУлBRҪбәПЈ¬ІўТэЖрПё°ыДЪ·ЦЧУҪб№№·ўЙъұд»ҜЈ¬Н¬КұХРДјПё°ыДӨЙПөДКЬМеёҙәПОпө°°ЧBAK1№ІН¬РОіЙBRI1-BR-BAK1КЬМеёҙәПОпЈ¬јӨ»оПё°ыДЪРЕәЕҙ«өјНҫҫ¶ЎЈ
ЎЎЎЎЈЁ3Ј©Пё°ыЦКBRРЕәЕј¶БӘ·ҙУҰЎЈЛжЧЕКЬМеёҙәПОпөДјӨ»оЈ¬BRРЕәЕҙУПё°ыНвҙ«ИлПё°ыДЪЎЈКЧПИЈ¬»о»ҜөДBRI1-BR-BAK1КЬМеёҙәПОпНЁ№эТ»ПөБРБЧЛб»ҜЧчУГјӨ»ојӨГёө°°ЧBSK1ј°ө°°ЧБЧЛбГёBSU1ЎЈЖдҙОЈ¬BSU1К№BRёәөчҝШТтЧУBIN2ТтИҘБЧЛб»Ҝ¶шөјЦВК§»оЈ¬ҙУ¶шКН·ЕКЬBIN2ТЦЦЖөДЧӘВјТтЧУBES1әНBZR1Ј¬К№Ждұ»БЧЛбГёPP2AИҘБЧЛб»о»ҜәуҪшИлПё°ыәЛЈ¬¶ФЬҝМҰЛШДЪхҘПмУҰ»щТтҪшРРЧӘВјЛ®ЖҪөчҝШЎЈ
ЎЎЎЎЈЁ4Ј©Пё°ыәЛBRПмУҰ»щТтЧӘВјөчҝШЎЈBES1әНBZR1КЗІОУлBRРЕәЕҙ«өјНҫҫ¶өДЦчТӘЧӘВјТтЧУЈ¬ҝЙұ»BIN2БЧЛб»ҜК§»оЈ¬¶шұ»PP2AИҘБЧЛб»ҜјӨ»оЎЈ»оРФBES1әНBZR1ФЪПё°ыәЛ¶ФBRПмУҰ»щТтұнҙпҪшРРЧӘВјөчҝШЎЈНЁ№эChIP-seqјјКхҝЙЦӘЈ¬ДвДПҪжЦРҙуФјУР5 000Ў«8 000ёц»щТтКЬөҪBRПа№ШЧӘВјТтЧУөДөчҝШЈ¬ЙПөч»щТтәНПВөч»щТтҙуФјёчХјТ»°лЎЈBES1әНBZR1ҝЙРОіЙН¬Фҙ¶юҫЫМеҪбәПФЪЛіКҪЧчУГФӘјюE-boxЈЁCANNTGЈ©ЙПјӨ»оBRЙПөч»щТтұнҙпЈ¬»тУлЖдЛыЧӘВјТтЧУРОіЙТмФҙ¶юҫЫМеҪбәПФЪЖф¶ҜЧУЙПBRREЈЁCGTG(T/C)GЈ©ФӘјюТЦЦЖBRПВөч»щТтұнҙпЎЈ
ЎЎЎЎBES1әНBZR1ҝЙЦұҪУУХөјЧӘВјТтЧУ№ІН¬өчҝШBRПмУҰ»щТтұнҙпЎЈАэИзЈ¬MYB30әНMYCАаЧӘВјТтЧУBIM1өДұнҙпКЬBES1УХөјЈ¬УЦҝЙәНBES1ө°°ЧҪбәП№ІН¬УХөјПВУО»щТтұнҙпЈ»¶шMYBL2әНHAT1УЦҝЙУлBES1№ІН¬өчҪЪBRПВөч»щТтұнҙпЎЈБнНвЈ¬BES1әНBZR1»№ҝЙНЁ№эХРДјTPLЧӘВјТЦЦЖёҙәПОпАҙКөПЦ°Р»щТтөДЧӘВјТЦЦЖЎЈ
3 ЬҝМҰЛШДЪхҘФЪЕ©ТөЙПөДУҰУГ
ЎЎЎЎЬҝМҰЛШДЪхҘЙъОп»оРФөДҫӯөдКөСйКЗҙу¶№ҪЪјдЙміӨәНЛ®өҫХнТ¶ЧӘҪЗКөСйЈ¬ТтҙЛЈ¬ЬҝМҰЛШДЪхҘКЧТӘөДЙъАн№ҰДЬКЗҙЩҪшПё°ыЙміӨЎЈЛжәуЈ¬ҝЖС§јТУЦІ»¶П·ўПЦЬҝМҰЛШДЪхҘёь¶аөДЙъАн№ҰДЬЈәҙЩҪшПё°ыЕтХНЎўПё°ы·ЦБСЎўЙъЦіЖч№ЩәНО¬№Ь·ўУэЎўПё°ыДӨј«»ҜУлЦКЧУұГЎўФҙҝв№ШПөј°РІЖИөчҪЪөИ·ҪГжЈ¬Н¬КұЈ¬ЬҝМҰЛШДЪхҘ»№ҝЙТФУл»·ҫіРЕәЕҪшРРҪ»»ҘЧчУГЈ¬У°ПмАҘіжәНХжҫъ·ўУэЎЈ
ЎЎЎЎЬҝМҰЛШДЪхҘәПіЙәНРЕәЕҙ«өјНҫҫ¶ФЪёчёцОпЦЦЦР¶јКЗұЈКШөДЈ¬ТтҙЛФЪЕ©ТөЙъІъЦРЈ¬ҝЙАыУГЬҝМҰЛШДЪхҘөДХвР©ЙъОпС§МШРФөчҪЪЕ©ЧчОпөДЙъіӨ·ўУэТФҙпөҪМбёЯЧчОпІъБҝәНЖ·ЦКөДДҝөДЎЈАэИзЈ¬АыУГНвК©ЬҝМҰЛШДЪхҘЈ¬өчҪЪКчДҫИНЖӨІҝ·ўУэЈ¬ТФҙЩҪшДҫІДәНПЛО¬ІъОпЙъіЙЈ»НЁ№эҝШЦЖBRРЕәЕНҫҫ¶Ј¬ҝШЦЖГЮ»ЁПЛО¬·ўУэЈ»ЕзИчЬҝМҰЛШДЪхҘЈ¬ҙЩҪш»Ё·Ы·ўУэј°КЪ·ЫВКЈ¬МбёЯ№ыКчЧш№ыВКЈ¬јхЙЩ»ыРО№ыРОіЙЈ»АыУГЬҝМҰЛШДЪхҘЈ¬өчҪЪ№ИОпМЗ·ЦЧӘ»ҜҙЩҪш№аҪ¬әНМбёЯЛ®№ыҝЙИЬРФМЗә¬БҝЎЈ
ЎЎЎЎЛжЧЕ¶ФЬҝМҰЛШДЪхҘРЕәЕПмУҰНҫҫ¶өДЙоИлБЛҪвЈ¬ОҙАҙОТГЗІ»ҪцҝЙТФНЁ№эНвФҙК©јУЬҝМҰЛШДЪхҘАҙКөПЦҙЩҪшЕ©ТөФцІъМбЦКөДДҝөДЈ¬¶шЗТҝЙТФНЁ№э·ЦЧУУэЦЦј°»щТтұајӯУэЦЦјјКхЈ¬К№ЬҝМҰЛШДЪхҘРЕәЕНЁВ··ЦКұҝХұнҙпәНЧйЦҜМШТмРФұнҙпЈ¬КөПЦЧчОпФЪЙъіӨҙЩҪшәНРІЖИДНКЬЦ®јдөДҫ«ЧјөчҪЪЎЈ ЈЁАҙФҙЈәЙъОпЕ©Т©КУҪзЈ©
Е©Т©ҝмС¶, 2020 (8): 50-51.


|






















